



Trophobioses between ants and hemipterans (PDF)
by Dirk Mezger, antbase.net
Ants are by far the most dominant arthropods in tropical rainforests in terms of biomass and abundance, and they govern key functions in their ecosystems (Beattie, 1985; Davidson 1997). Most of them are omnivore because for a solely predacious life style, there is not enough prey. Many of them depend on trophobioses with hemipterans. Ants gather sugar and amino acid containing exudates from these plant sap sucking insects. This can be an advantage for both ants and hemipterans because the hemipterans are protected against predators and parasitoids, and mould is less likely when the sugar containing exudates are removed by ants. Even the plants can benefit from these trophobioses when herbivorous insects are removed by ants. |
 |
|
| Fig. 1 Coreid bug associated with Crematogaster modiglianii on climbing bamboo (Dinochloa trichogona) | Fig. 2 Delphacids associated with Lophomyrmex cf. bedoti on climbing bamboo (D. trichogona) |
Climbing plants like Spatholobus are also important host plants for some hemipterans visited by ants. Especially some species of cicadellids and membracids can be found on these plants. In a few cases there are visited by Crematogaster modiglianii and Camponotus rufifemur at the same time. This ant species live together in so called parabioses (Menzel et al. in prep.). That means that these ant species share a nest in a tree hole.
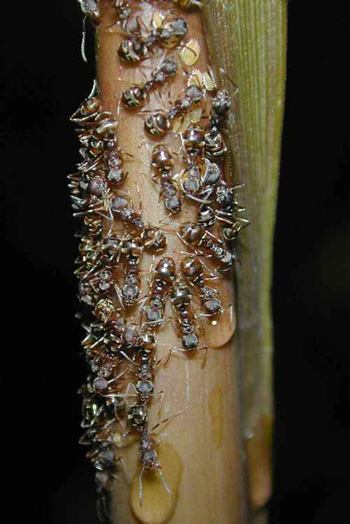
On seedlings of large trees like Parashorea (Dipterocarpaceae), trophobioses can be found often (fig. 3). For these seedlings the trophopbioses are especially important because they seem to be vulnerable to herbivores and the ants can protect them.
Most of the trophobioses
are found in the canopy of the large trees but they are out of sight.
Observing the large trails of ants along stems of the large trees one
can assume that there a lots of plant sup sucking insects on the tree.
|
 |
|
| Fig 3 Larval stages of an unidentified cicadellid together with Lophomyrmex cf. bedoti on a Parashorea seedling |
 |
Dolichoderus herdsmen ants are only found in South East Asia. These ants have a very close association with their hemipteran partner (Dill et al. 2002). These ants are associated with a special group of mealy bugs (Allomyrmococini). The mealy bugs are continuously guarded, brought to the biwak nest of these ants at night time and carried to their host plants which include a broad array of plants including the climbing bamboo (fig. 4) mentioned above. In order to transport their hemipteran partner to suitable host plants these ants have a nomadic lifestyle (Maschwitz & Hänel 1985). These associations is very specific, every of these ant species has its own mealy bug species. |
| Fig. 4 Dolichderus maschwitzi (herdsmen ants) together with Promyrmococcus on D. trichogona |
When leaving the forest and looking to areas covered with low secondary vegetation, there is less diversity of hemipterans. Many areas are covered by the invasive weed Chromolema odoratum. On these plant originally from Central America, only two species of aphid (Aphis spinocola and A. gossypi), and sometimes one species of mealy bug and a coccid can be found. When these areas are not too far away from the forest, quite a lot of ants from all subfamilies can be found on these associations if no aggressive invasive ants are present. When these invasive species like e.g. Anoplolepis longipes have already occupied the site, only few other ant species visit trophobioses nearby them.
Literature
Back to the Ecology overview